
Eureka, today’s post covers some general information about the morphology of bacteria, yeasts and other microorganism on agar plates and why it is important to know at least a bit about it to get the most information out of your agar platings.
Q: What do you mean by morphology of colonies?
The morphology of a colony describes how microorganisms appear on agar media such as Sabouraud, malt agar etc. Morphology just describes the colonies. If you streak some microorganisms on agar plates, they grown (if the media is appropriate for this particular organism) and form visible colonies. The colonies appear as spots like shown in Fig 1. It is important to remember that a colony are thousands to millions of microorganisms together, not a single microorganism cell. Ideally all the cells within a colony originated from one single cell at the beginning (clonal expansion). If single cells are closer together on the agar, the individual colonies overlap and no single colonies are visible (left-upper part in Fig 1). In this case, the concentration of the yeasts is just too high to observe individual colonies.

Fig 1: Brettanomyces bruxellensis on Sabouraud agar plate after 11 days
Fig 1 shows what you get if you streak some Brettanomyces yeast on Sabouraud agar. The roundish spots are the colonies (as you can see on the right side in Fig 1).
Q: How do you get single colonies?
To get an accurate description of a colony, single colonies are necessary. But how do you get single colonies in the first place? As mentioned above, if the individual cells after streaking are to close to each other, the colonies might overlap. To get single colonies one simply has to ensure a low concentration to prevent such colony-overlays. One example to do so is to dilute the cells directly on the plate itself by using a special streak technique called dilution streak or Z-streak (Fig 2). How this is done is shown in a video (YouTube) as well.

Fig 2: Dilution streak done with three streaks
Begin with a cell suspension. You might even use a yeast slurry in the first place. A first streak is done to get some cells on the plate (Fig 2, streak 1). One expects a lot of cells visible on the trajectory of the first streak and the individual colonies overlay each others. After the first streak, you sterilize your inoculation loop, let it cool down and collect some cells by passing the inoculation loop through the first streak for a second one. This time the concentration of cells is already lower because you only pick a subset of yeast cells. This process can be done for a second time to get three streaks in the end (Fig 2). The plate after a dilution streak might look like shown in Fig 1. Unfortunately, there are no colonies visible in the third streak anymore. Anyway, I hope you get the idea.
Single colonies are not only useful to describe their morphology but also to differentiate between different microorganisms. For instance, if you are interested in separating the Saccharomyces yeasts (brewer’s yeast) from Brettanomyces yeasts you can use the dilution streak and hopefully some colonies arise from single Saccharomyces colonies and others from Brettanomyces cells.
Q: Why is the morphology important?
Lets assume the morphology of a colony, representing one kind of microorganism (remember the concept of the single cell at the beginning), is unique for every microorganism there exists. The morphological description could therefore be used to identify the kind of microorganism on your agar plate. This is just an assumption because there are a lot of microorganisms which have similar morphologies. To summarize, the morphology of the colonies can be useful to identify the kind of microorganism you have on your plate. Lets go through some examples. Have a look at Fig 3.

Fig 3: Girardin bugs on Sabouraud agar plate
I assume it is obvious that there are different kinds of colonies and hence morphologies. There are differences in shapes, size and colors. To conclude, different morphologies originate from different microorganisms. I can give you even further information here. The white colonies (big ones and wavy) are yeast cells, the flat beige ones bacteria. The very small white colonies are another kind of bacteria. You see, the morphology can even be used to differ between yeasts and bacteria. That’s why agar media are very common in microbiology labs to identify different kinds of yeast/bacteria. One application here could be to test a beer for spoilage organisms such as Lactobacillus (beer turned sour). Plate some of the sour beer on a plate where Lactobacillus can grow and if colonies arise with a typical Lactobacillus morphology, you can be certain to have a Lactobacillus contamination in your beer. I will not get into further detail about the different media and strategies used to do these tricks. Just to give you an idea what the whole agar media method is capable of.

Fig 4: Water kefir on Sabouraud agar plate
Maybe an example to show that the colonies are not always circular. Some microorganisms tend to form large flat colonies as it can be seen in Fig 4. In this case, I plated some of my kefir culture on a Sabouraud agar plate. You can even observe some yellowish colonies. Colonies are not always white or beige either. Not only can you choose different kind of agar media but also add some dyes for further characterization. One such example is shown in Fig 5. In this case bromocresol green is added to differentiate between microorganisms that can grow as white colonies and such as green ones. The color differences suggest that there are at least two different kinds of microorganisms on the plate shown in Fig 5.

Fig 5: Jolly Pumpkin’s Madrugada Obscura dregs on bromocresol green Sabouraud agar
Q: How do you determine the morphology of a colony?
First you need a pure culture of the microorganism. This is important because the morphology can differ if other microorganisms are in the same colony. The morphology can even be different on other agar media. Lets assume you want to describe the morphology of a pure brewers yeast (Saccharomyces cerevisiae). The first thing to do is streaking the yeast on a suitable agar media with a dilution streak and incubate the plate until colonies arise like shown in Fig 6. Sabouraud is a typical agar media for Saccharomyces and other yeasts. Malt agar media works as well.

Fig 6: Wyeast’s 2112 California Lager on Sabouraud Agar plate
In case of Fig 6, I streaked some of Wyeast’s 2112 California Lager yeast on a plate to check the purity. Now what about the morphology? Lets take a single colony and describe the following characteristics: form, margin, elevation (shape of the colony from the side), size, texture, appearance, pigmentation, opacity. The following descriptions are just an example.

Fig 7: from: http://commons.wikimedia.org/wiki/File:Bacterial_colony_morphology.png#filelinks; (Adapted and redrawn from Seeley, HW & Vandemark, PJ (1962) Microbes In Action: A laboratory manual of microbiology. WH Freeman (San Francisco, London) by user Ewen)
One might describe the colonies shown in Fig 6 as following:
Margins: Entire
Form: Circular
Elevation: Convex
Surface: Smooth
Opacity: Not transparent, shiny
Color: Off-white
That is what you can expect when you streak a yeast colony on a Sabouraud plate. The morphology of Saccharomyces is very similar on malt agar. Maybe some of you observed that there are yet some other different colonies on the plate in Fig 6. There were some impurities in this yeast sample as expected in the first place.
Q: Is the morphology of a given microorganism always the same?
Unfortunately not. The morphology of colonies can depend on the type of agar media used, if oxygen is present, nutrients, vitality, pH-levels, incubation time, other microorganisms present… Just keep in mind that a morphology description is not universal. If you encounter a morphology description of a specific microorganism, always check the type of agar media used and the conditions how the plates were incubated.
Q: Is it possible to differentiate between Saccharomyces and Brettanomyces based on morphology?
One of the most simple tricks to differentiate between the two yeasts is the incubation time. Saccharomyces colonies arise relatively quickly (within few days). Brettanomyces grow much slower (days to weeks). The second trick is to use a microscope and have a look at the different colonies. A third one might be (haven’t tried that one yet) to inhibit the growth of Saccharomyces by adding some growth inhibiting substances. Differentiating those two yeasts based on morphology is not that easy in my opinion.
Q: Is it possible to differentiate between top and bottom fermenting yeasts or even yeast strains based on colony morphologies?
As far as I know and from my experiences, differentiating between bottom and top fermenting yeasts base on colony morphologies is not possible. And it is not possible as well to differentiate between different yeast strains as well. Although I encountered some different morphologies for wheat strains at one point. However, I would not do any strain differentiation based on morphologies.
I hope there were some useful information in this post to give you a better understanding of agar media cultivation. Agar media are a very powerful tool in microbiology and is also widely used in breweries to check for impurities in beer or water. Understanding the concept of colony morphologies is therefore very important to get the most information out of agar media cultivation. And know about some limitations of the method as well.



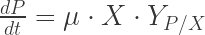









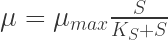
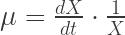










{kind=link}