
Eureka, second yeast kinetic post. I further discuss Monod growth kinetic models with the inclusion of some inhibition parameters. I would advice you to first read the yeast kinetic introduction post if you haven’t done so already. One inhibition phenomena has been studied on Brettanomyces by looking at growth behaviour under aerobic and anaerobic conditions and the influence of initial acetic acid. All the values of the coefficients are taken from a publication written by Yahara et al (2007). The goal of this publication was to investigate the glucose utilization rate of Brettanomyces bruxellensis at different acetic acid levels under aerobic and anaerobic conditions. The authors first conducted experiments and then proposed an extended Monod model to simulate the growth kinetics under different conditions.
I would like to start by discussing the basic equations used for the extended Monod model used by Yahara et al (2007). The basic Monod equations have been discussed in my introductory post.



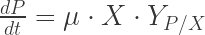
- µ specific growth rate [h-1]
- µmax maximum of the specific growth rate [h-1]
- X is the biomass concentration [g L-1]
- S is the substrate concentration [g L-1]
- KS is the substrate saturation constant [g L-1]
- Ki is the reciprocal of the inhibitor constant [L g-1]
- q is the exponent for the inhibitor constant and biomass [ – ]
- YX/S Biomass on substrate yield coefficient [gX gS-1]
- YP/X Product on biomass yield coefficient [gP gX-1]
- dX is the change of the biomass concentration [g L-1]
- dS is the change of the substrate concentration [g L-1]
- dP is the change of the product concentration [g L-1]
- dt is the change of time when dX and dS happen [h]
The first differential equation for the specific growth rate µ is now slightly modified to include an inhibition factor Ki and an exponent q. The authors cultivated Brettanomyces bruxellensis under aerobic and anaerobic conditions with varying initial acetic acid concentrations (1, 2, 3 and 4 g L-1). The substrate (glucose) and inoculation rate were the same throughout the whole experiments. Two products were included into the model, ethanol (P1) and acetic acid (P2). The only difference in the Monod model here are two different yield factors (YP/X) and one equation for ethanol (dP1/dt) and one for acetic acid (dP2/dt). µmax was obtained the same way as I showed in the introduction post. All the remaining coefficients were obtained by iterative approaches.
I would like to show some of the growth curves published by Yahara et al (2007) which I obtained by using their coefficient values running the model using Berkeley Madonna.
Aerobic growth and acetic acid concentrations
Aerobic growth and different initial acetic acid concentrations. All the graphs show the substrate concentration (glucose) in red, the biomass concentration in black, the ethanol concentration in green and the acetic acid concentration in blue. In addition, I included the values of the coefficients in the individual graphs.

Fig 1: Aerobic growth with initial 1 g L-1 acetic acid. Glucose (red) g L-1, yeast biomass (black) g L-1, ethanol (green) g L-1, acetic acid (blue) g L-1
The first graph shows the growth one can observe under aerobic conditions and an initial acetic concentration of roughly 1 g L-1 (Fig 1). The glucose is fully metabolized by the yeasts within 100 h of cultivation. It can also be observed that Brettanomyces produce ethanol and some acetic acid.

Fig 2: Aerobic growth with initial 4 g L-1 acetic acid. Glucose (red) g L-1, yeast biomass (black) g L-1, ethanol (green) g L-1, acetic acid (blue) g L-1
The next graph shows the growth under aerobic conditions and an initial acetic concentration of roughly 4 g L-1 (Fig 2). In this case the glucose is not fully metabolized after 100 h as previously shown (Fig 1). The Brettanomyces still grow but at a slower rate. Still some ethanol is produced and a minor amount of acetic acid.
From these two graphs one can already conclude, that the amount of initial acetic acid in the media seems to significantly impair the growth of Brettanomyces. At higher acetic acid levels the Brettanomyces seem to grow substantially slower.
Anaerobic growth and acetic acid concentrations
The next graphs show the growth curves in absence of oxygen again with different initial amounts of acetic acid.

Fig 3: Anaerobic growth with initial 1 g L-1 acetic acid. Glucose (red) g L-1, yeast biomass (black) g L-1, ethanol (green)
The graph shows the growth one can observe under anaerobic conditions and an initial acetic concentration of about 1 g L-1 (Fig 3). Yet again the glucose is fully metabolized within 140 h of cultivation as previously observed under aerobic conditions and low initial acetic acid concentration (Fig 1). Although the Brettanomyces under anaerobic conditions seem to metabolize glucose at a slower rate than under aerobic conditions. The Brettanomyces produce again ethanol. But no measurable amount of acetic acid. Under anaerobic conditions, Brettanomyces produces much more ethanol. In comparison to aerobic condition, the Brettanomyces grow faster in presence of oxygen.

Fig 4: Anaerobic growth with initial 6 g L-1 acetic acid. Glucose (red) g L-1, yeast biomass (black) g L-1, ethanol (green) g L-1
At higher initial acetic acid concentrations and anaerobic conditions, Brettanomyces still grow but again at a slower rate (Fig 4). A lot of the glucose is not metabolized after 140 h of cultivation. The yeasts still produce some ethanol but the growth curve of the biomass stays roughly the same. Indicating a very slow growth rate.
Because the authors could not measure any acetic acid production under anaerobic conditions, one can conclude that the yeasts do not produce measurable amounts of acetic acid under anaerobic conditions. In addition, higher levels of acetic acid inhibit the growth of the yeasts.
Summary
Results published by Yahara et al (2007) showed:
- B. bruxellensis grows faster in presence of oxygen
- B. bruxellensis produces ethanol under aerobic and anaerobic conditions
- More ethanol is formed under anaerobic conditions
- In presence of oxygen and low amounts of initial acetic acid, B. bruxellensis can produce up to 4 g L-1 of acetic acid
- Acetic acid can inhibit the growth of B. bruxellensis
- No measurable amount of acetic acid is produced under anaerobic conditions
- The proposed model based on a Monod model can describe the dynamic growth curves of B. bruxellensis
Uscanga et al (2003) already showed that B. bruxellensis grows faster in aerobic conditions, produces acetic acid in presence of oxygen, and higher initial amounts of acetic acid inhibits the growth of B. bruxellensis. In addition, Uscanga et al (2003) further showed that higher oxygen amounts lead to a decrease in glucose metabolization, the amount of ethanol produced decreases and the acetic acid level increases. This might be an indicator that high levels of oxygen inhibit the metabolism of B. bruxellensis as well like high initial acetic acid levels.
For the brewers: Brettanomyces slow grower under anaerobic conditions, form more ethanol but no acetic acid. Acetic acid is only produced if oxygen is present.
Quick word about models
One has to keep in mind that this model can’t describe everything. For example, if one runs the model using a high substrate concentration, the Brettanomyces will continue to grow even in presence of very high ethanol concentrations. In reality, Brettanomyces have an alcohol tolerance as well where they stop growing. However, this can easily be included in the model. This example was just to show that one has to be careful with models.
I hope this post was interesting to read and gave you an idea how models can be used to describe growth behaviours under inhibitory conditions.
The next post about yeast kinetic models will be concerning yeast calculators.
Bibliography
- Kurtzman CP, Fell JW, Boekhout T (2011) The Yeasts, a Taxonomic Study. Volume 1. Fifth edition. Elsevier (Link to sciencedirect)
- Uscanga MG, Délia ML, Strehaiano P (2003) Brettanomyces bruxellensis effect of oxygen on growth and acetic acid production. Appl Microbiol Biotechnol. 60: 157- 162; DOI: 10.1007/s00253-002-1197-z
- Yahara GA, Javier MA, Tulio MJM , Javier GR, Guadalupe AUM (2007) Modeling of yeast Brettanomyces bruxellensis growth at different acetic acid concentrations under aerobic and anaerobic conditions. Bioprocess Biosyst Eng. 30: 389 – 395; DOI: 10.1007/s00449-007-0135-y

Pingback: 80 – Funky Maple 90 Minute Clone | Funky Horse Brewer
Hello,
I like this idea of a kinetic model for yeast growth, and I am trying to do something similar for cider making.
However, trying to figure how the extended Monod equation works, I think you might have done a transcription error because the units are not consistent as it is currently written…
you have: MU = MUmax [S.X / (denominator)]
Since MU have the same units as MUmax, the term in brackets should be non-dimensional (as in the basic Monod equation), which is not the case here.
I haven’t read the Yahara paper, but my guess is that the X term in the numerator has no business there, and that the units of the K_i term in the denominator should be L/g instead of g/L.
This way, all units would be consistent and the term in brackets would be non-dimensional.
Thanks for your attention,
Claude
Hi Claude,
you are absolutely right. Removed the X term and the unit of K_i is indeed L g-1. Thanks for your corrections and reading the post very carefully.
Cheers, Sam